Evolution & Systematics
Fossil evidence of sipunculids may not be as extensive as that of other hard-shelled calcareous species, but this evidence suggests a common sipunculan-molluscan ancestor existed in the Ediacarian. In addition, studies suggest sipunculids were living a benthic existence as early as the Cambrian, and burrowing sipunculids – such as P. nigrescens – have been found associated with corals since the Mid-Paleozoic (Cutler, 1994). This was suggested by Brett & Cottrell (1982) who found rock boring sipunculid fossils inside the Devonian tabulate coral Pleurodictyum. Likewise, trace fossils of sipunculid burrows from the early Ordovician, have been identified (Brett et al., 1983; Pemberton et al., 1980). The only legitimately unambiguous fossil record of a sipunculid was uncovered in southwest China from the Cambrian era (Huang et al., 2004). Ottoia prolifica (Figure 16) has been identified as the best candidate for a sipunculid ancestor among the Burgess shale fossils (Banta & Rice, 1976).
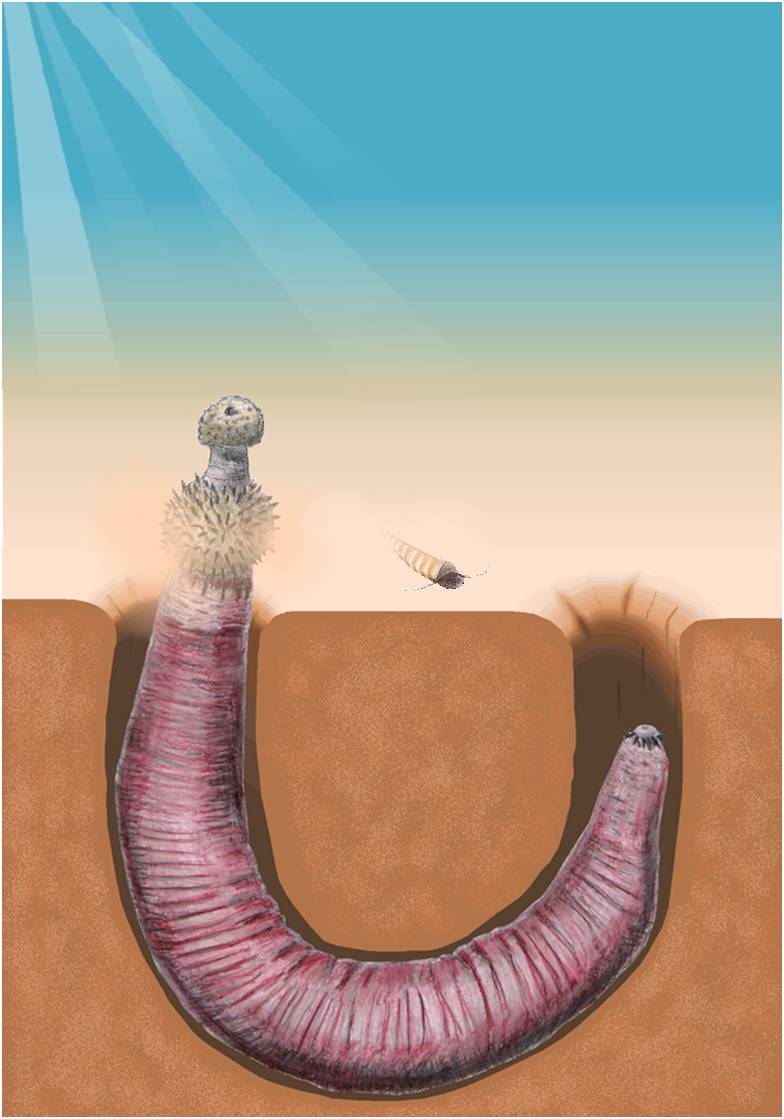 Figure 16: Ottaia prolifica burrowing in sediment, reproduced from Wikimedia under the Creative Commons Attribution-Share Alike 3.0 Unported licence.
Sipunculan phylogenic position has remained a contentious debate between taxonomists. Morphologically, they strongly resemble Annelids without segmentation. General agreement is that Sipuncuids are protostomes and should be classed under the Lophotrochozoa, bearing to similarities with Molluscs and Annelids (Cutler, 1994; Schulze et al., 2005; Saiz, 1993). Studies done on embryological similarities between Sipunculids, Molluscs, and Annelids have all been ambiguous. The trocophore larva present in sipunculids seems to sugest affinities with molluscs, and annelids, and the molluscan cross during cleavage seems to suggest a closer affinity to molluscs (Ruppert et al., 2004). However recent studies suggest the use of molluscan and annelidan crosses as phylogenetic tools may be of limited importance (Schulze et al., 2005).
Similarities to molluscs include homologies in the head regions of larvae, similarities between the molluscan foot and the sipunculan lip. Studies by Scheltema (1993) suggest the sipunculan lip glands may also be homologous to molluscan pedal glands, and also suggest the sipunculan buccal organ to be of similar origin as the molluscan radular sac. On the other hand, similarities to annelids include the distinctive similarities between prototrochal and metatrochal bands in the larvae of both (Rice, 1985). Likewise, the egg envelope of some species is retained to form the larval cuticle. In Phascolosoma agassizii a close relative or P. nigrescens, the nerve cord is doubled in early larval stages, as in many Annelids (Rice, 1985).
Cladistic analyses of morphological, molecular, and gene order data have provided many hypotheses relating Sipuncula to many unresolved taxa (Schulze et al., 2005). Little agreement is found throughout most of these, and the exact classification of Sipuncula remains elusive. Over the years, the taxa ‘Sipuncula’ has even changed taxonomic level, and existed as a family, order, sub-class, and even a class (Schulze et al., 2005).
A combined analysis of morphological as well as genetic, and molecular data was undertaken by Schulze and colleagues (2005) and rendered the following consensus tree (Figure 17).
Figure 17: Strict consensus tree of the three most parsimonious trees generated by Schulze et al (2005). Analysis was based on combination of 18S RNA, 28S rRNA, histone H3, and morphological data. White shapes represent Sipunculidea, white circles are Sipunculidae, white stare are Phascolionidae, white squares are Thermistidae, and white triangles are Golfingiidae. Black shapes represent Phascolosomatidae, black circles are Asidosiphonidae, and black squares are Phascolosomatidae. |